

Of all the powers of the animal world that humans have envied and engineered into submission, nothing has become a more overdone trope of this desire than the flight of birds. In our endeavor to understand flight, we have broken down the components of aerodynamics into something we can quantify, calculate, and model. This approach has obviously worked quite well for us. Just think about how a Boeing 747 jumbo jet can haul passengers from the East to West Coast of America in about six hours. The same journey would have taken wagon-driving pioneers months of life-threatening travel through forests, endless plains, deserts, snowy mountains, more deserts, and more mountains.

With our highly engineered view of flight, we tend to look back at living organisms and try to find elements of evolutionary design that seem perfectly adapted to certain modes of life. For example, let’s take a look at the nostrils of two very different birds, the Peregrine Falcon (Falco peregrinus) and gannets (Family Sulidae).
The Peregrine Falcon is a small North American raptor famous for its incredible high-speed dives. These dives, or stoops, are generally reported to clock in at 200 mph (320 km/h). This makes the Peregrine one of the fastest animals on Earth, “a feathered bullet dropping out of the sky (Hagler 2012).” Traveling at such speeds requires many modifications, including some less obvious ones like redirecting airflow into the nostrils for breathing. It is repeatedly said that the force of air entering the nose at 200 mph would cause the lungs to explode. In order to prevent this from happening, there are bony tubercles in the nares that act as baffles to safely regulate the passage of air into the respiratory system. As a matter of fact — according to these often repeated anecdotes — the nostrils of Peregrine Falcons inspired the design of inlet cones for supersonic jet engines.
That’s a cool fact, but what does it really mean?

To begin with, traveling at high speeds will not cause lungs to explode. Quite the opposite. There are two physical principles to keep in mind here: (1) the Bernoulli Effect where higher air speed results in lowered air pressure and (2) the energetically favored direction is always from high to low. As the Peregrine Falcon reaches top speed during the stoop, the increasing air speed encountered by the nares will result in a drop in air pressure. Inhalation relies upon relatively high pressure air outside the body rushing into the low pressure area of the lungs. Eventually, the air pressure outside the stooping falcon will approach equilibrium with the air pressure inside the lungs, making breathing very difficult. Think about how it is harder to breathe when facing into a strong wind, sticking your head out a car window on a freeway (not necessarily recommended), or riding a fast boat. The presence of bony tubercles in the falcon’s nose act to slow down the airflow, increasing the air pressure, and allowing air to be drawn into the body. Seems like a clever bit of evolutionary adaptation to extreme high speed flight.

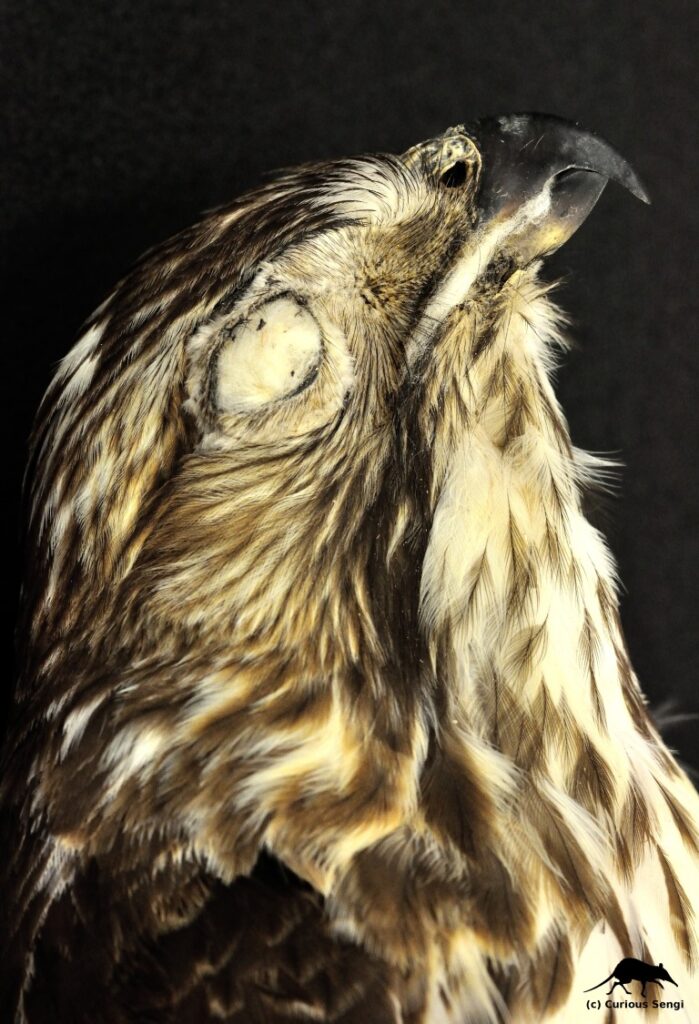
This tidy story is somewhat disrupted once we take a look at the phylogenetic distribution of narial bony tubercles. The Peregrine Falcon shares this morphological feature with all members of the Family Falconidae, which includes raptors of a wide variety of shapes, sizes, and flight abilities. At one end of the spectrum is the Peregrine Falcon that knocks its bird prey out of the air. At the other are the caracaras, which soar at a leisurely pace searching for carrion, much like vultures. This leaves us with a puzzle: if the tubercles within the nostrils of the Peregrine Falcon are adaptive for respiration at extreme flight speeds, why do all members of the Falconidae possess these tubercles? It is possible that narial tubercles were present in the common ancestor of all Falconidae and, therefore, all its descendants still carry this feature. Perhaps the tubercles evolved to serve a different function — such as sensing airspeed or temperature — and this existing structure was modified with a new purpose in the Peregrines. The conclusion is that we do not fully understand why the bony tubercles of the nostril appear in the Falconidae and what adaptive purpose (if any) it may serve.
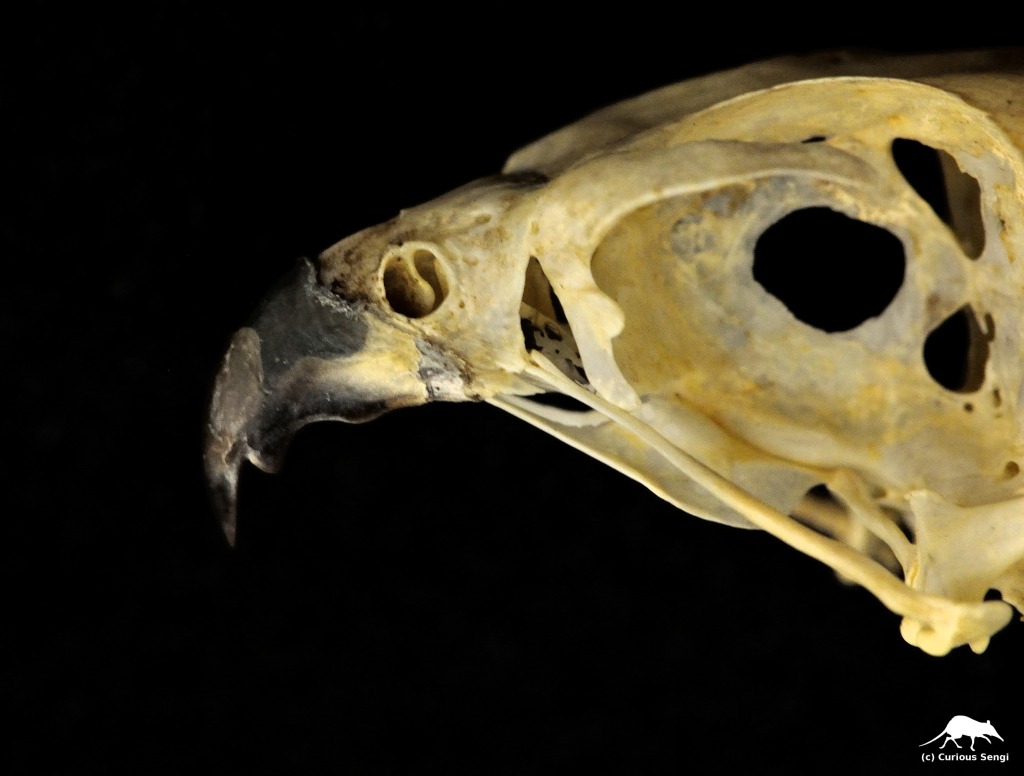

Perhaps we felt confident in stating the purpose of the Peregrine Falcon’s unusual nostrils because we saw how engineers solved the problem of regulating air intake in jet engines in a very similar way. In the post-WWII years, military aircraft were breaking more speed records with an ever sophisticated understanding and use of rocketry, but aircraft could only travel so fast until the engines would choke and then stall. It was soon discovered that instead of passing through the cylinder of the jet engine, air flow was being diverted away, taking with it the oxygen necessary for combustion. This is the same problem caused by the Bernoulli Effect and the flow from high to low pressure discussed earlier. The addition of cone-shaped structures in the engine’s inlet generates shockwaves that slow down airflow and allow the engines to continue running. The inlet cone innovation made supersonic flight possible. In 1947, Chuck Yeager was able to take a Bell X-1 experimental plane faster than the speed of sound, i.e., Mach 1, which is a blinding 768 mph (1235 km/h) at sea level.
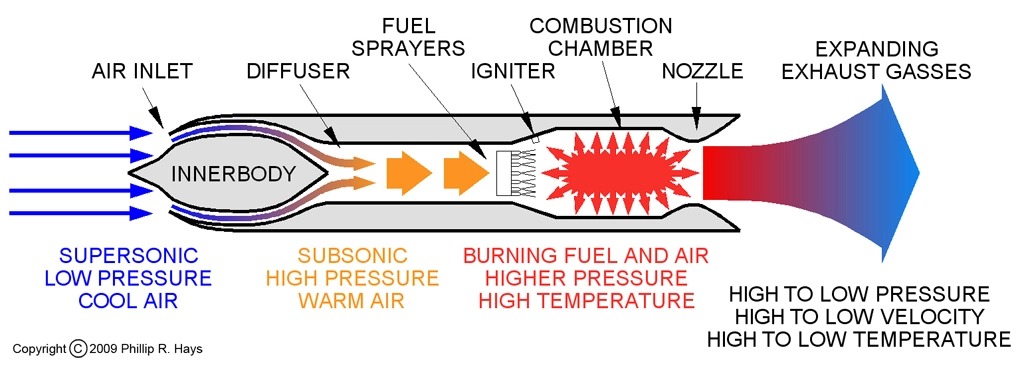
The similarity between the design of the inlet cone in supersonic jet engines and the bony tubercle in the nostrils of Peregrine Falcons make for another tidy story where Nature directly informed engineering. Though I was unable to find any literature that proved experimental research was done on the aerodynamics of falcon nostrils, it does not preclude the possibility that a casual observation inspired an idea.

Another fast flyer encountering extreme physical forces are gannets. These seabirds have been observed plunge diving from a height of about 100 feet (30 m), drawing their wings back and configuring their bodies into a tight, streamlined shape to pierce the surface of the water where they capture fish. At the moment of impact with the water, gannets can be traveling at 54 mph (86.4 km/h, or 24 m/s). (There are macabre data collected from 169 suicides of people jumping off the Golden Gate Bridge in San Francisco. Impact velocity was calculated to be approximately 33 m/s. Almost 100% of jumps were fatal, with a vast majority of deaths caused by the impact itself.) Like the Peregrine Falcon, gannets have evolved a suite of features that allow them to cope with such intense hunting strategies. As many human swimmers have experienced, how does the gannet dive without getting water up the nose?

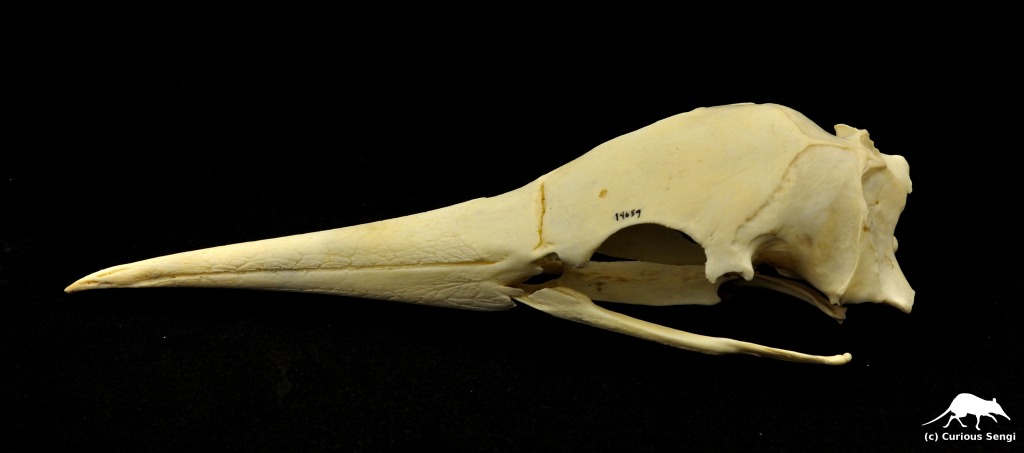
Gannets are believed to bypass the problem by losing the external nares entirely.
As embryos in the egg, gannet nostrils develop identically to many other bird species, with the nostril openings and immediate vestibular cavity sealed by a plug of epithelial tissue. While this plug breaks down a little bit later in development to open up the nares, it remains in gannets. Eventually, the gannets’ external nares are overgrown with bone and covered by the keratinous sheath of the beak, the rhamphotheca. Interestingly, while the nostrils are completely occluded and there is no flow of air through the nasal cavity, gannets still retain well-developed olfactory structures.


How do gannets manage without nostrils? Macdonald (1960) described secondary external nares — compensatory nostrils, if you will — formed by a gap at the corner of the mouth where the upper beak overhangs the lower. This area of overhang is made from a bone called the jugal (equivalent to our cheekbone), which is covered by a hinged plate of keratin. These two elements are loosely connected to the rest of the skull and are likely to collapse against the sides of the beak from the external pressure of water when diving, thus passively closing up these secondary external nares.

The total loss of nostrils in gannets seems to be a great way to prevent water from forcibly entering the nose during plunge diving and potentially causing damage or water entering the respiratory system. However, Macdonald noticed some interesting patterns in other unrelated diving birds. Cormorants (Family Phalacrocoracidae) certainly dive, but from the water’s surface, not from the air. Despite this more gentle entry into the water, cormorant nostrils are small and almost completely occluded. In contrast, the Brown Pelican (Pelecanus occidentalis) is a heavy-bodied plunge diver with open nostrils. The nostrils are surrounded by a flap of skin that might push up against the nostril and seal it like a valve under external water pressure.
To say that the complete narial occlusion in gannets is directly correlated to plunge diving would be ignoring some of the complexities of the story. We could interpret the near-total loss of nostrils in the cormorant as a remnant from a plunge diving ancestor. Or it could evolved for a different reason entirely. The alternative mechanism for closing up the nares in the Brown Pelican is suggestive that there is an adaptive advantage to not getting water up your nose when diving. In addition, the employment of a different solution to the same problem suggests that there are negative trade-offs involved with losing nostrils. For example, seabirds need salt-secreting glands to rid their bodies of the excess salt they ingest. These glands usually empty out through the nostrils. Gannets have relatively small salt-secreting glands and must discard concentrated salts from their mouths.

Interpreting the function of anatomical structures is always going to be more nuanced than saying “structure X is perfectly adapted to serve purpose Y.” The biological world is far removed from the world of engineering design. In our interpretations, we have to take into account the baggage and constraints imposed by evolutionary history, experimentally prove that a given structure does have a function that enhances performance, and keep in mind that trade-offs exist.
Leave a Reply